



27 Dec Cytoskeleton Components in Cognition
A Neuron’s Skeleton
Unlike the external structure of many spheroid and amorphous cells, the external structure of neurons is complex and rigidly determined. As you may remember from earlier posts, the many different types of neurons residing in particular cortical strata (in layers of the gray and white matter) each have distinctive characteristics that enable them to perform specialized functions and interact with other specific cell types. The extent of arborization of the different cell types structurally determines the number of synaptic junctions possible.

The structure of links between neurons is maintained by the cytoskeleton. Just as bones provide more than structural support (for example, bone marrow also provides red and white blood cells), cytoskeletal members serve additional roles in cells. Two of the known roles of cytoskeletal components include maintaining the link structure or the physical form of neurons and transporting cell components from the soma to the synapses. Possible cybernetic functions of the cytoskeleton will show that cells have complex mechanisms capable of storing data.
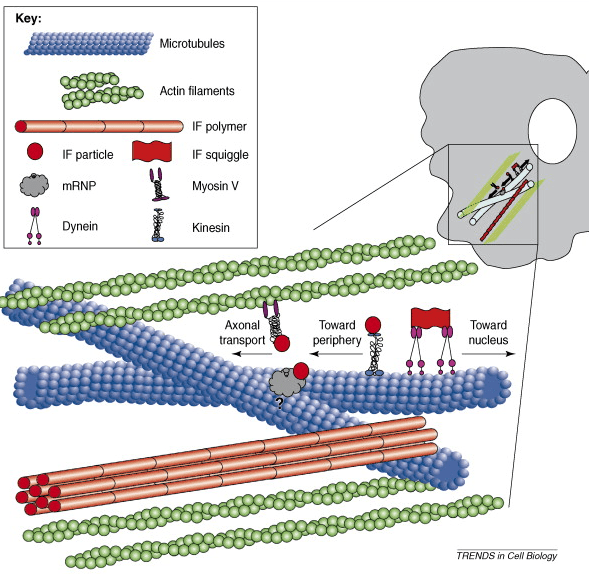
The illustration above shows that filaments are everywhere in neurons. Neurons possess far more filaments than can possibly be shown here. While some neural network literature suggests that neurons may average on the order of 10,000 connections each, upon closer scrutiny it appears that many or most neurons have less than 1000 synapses. The synapses of some may actually number only in the dozens.
| Understanding Context Cross-Reference |
|---|
| Click on these Links to other posts and glossary/bibliography references |
|
|
|
| Prior Post | Next Post |
| Myelin Sheath Function | Microtubules |
| Definitions | References |
| cytoskeleton | Hameroff & Watt 1982 |
| arborization | Kater & Letourneau 1985 |
| intermediate filaments (IF) | Steinert & Roop 1988 |
| electrical flow (action potential) | microtubules (MT) |
Cytoskeleton Components
The cytoskeleton is made up of protein-based structures called microfilaments (also known as actin filaments), intermediate filaments (IF), and microtubules. The filaments are hair-like strands. There are many of them in all parts of neurons except the nucleus (excluding lamins, a type of IF). As described earlier, the branching or arborization of neurons is extensive, rigidly determined, and critical to their functionality.
In addition to providing structural support, the cytoskeletal members may provide some of the information and locomotion necessary for growth. They propel their extending fibers to exactly the right place for them to fulfill their required functions [see Kater & Letourneau, 1985]. It has been suggested that information-processing functions are also performed by these filaments [see Hameroff & Watt, 1982].
Their possible roles in cognitive processes are due to the complex chemical structures of the filaments that permit them to respond differently to changes in the electrical potential within the cell’s cytoplasm or axoplasm.
Intermediate Filaments

IF are fibrous bodies that make up the cytoskeleton of most cells. Although there are five types of IF, only two are found in neurons: neurofilaments and lamins. The neurofilaments are ubiquitous outside the nucleus; lamins are found inside the nucleus. Together, they constitute strands of linked protein chains.
Although there are as many as 15 different proteins each in some types of IF (Types I and II), neurofilaments (Type IV) and lamins (Type V) contain only four proteins each [Steinert & Roop, 1988, p. 595]. The proteins in neurofilaments are classified as NF-L, NF-M and NF-H. An additional protein may be found in peripheral neurons. Neurofilaments (Type IV) are the subject of this post, but they are often referred to by the generic term IF.
(Actin filaments are too small to see.)NOTE: Microfilaments are the most numerous type of filament in neurons. In muscle tissue, microfilaments are integral to expansion and contraction. Their participation in cognitive processes has not yet been established.
Neurofilaments
Neurofilaments are the largest of the intermediate filaments. They range in size from 57-150 kd while IF found in most other cells are no larger than 70 kd. Neurons contain hundreds to thousands of neurofilaments each; these are located in the somas, the axons, and the dendrites. They are NOT found in the nucleus. Lamins (Type V) are the only IF found in the nucleus. Intermediate filaments are so designated because of their size. They fall between the smaller microfilaments composed of actin and the larger microtubules (MT) composed of tubulin. These three types of filaments make up the cytoskeleton of cells. Of the three, IF are the most structurally and functionally complex.
As with many of the illustrations, the representations in this post are not to scale in either size or quantity (of the neurofilaments, for example). The illustration above, however, gives you an idea of quantity. That illustration actually resembles photographic images of an axon section. The scale, too, is indicative of both the relative size and quantity of filaments in an axon. The size and the frequency of MT in the illustration above are also representative.
IF Structure
Intermediate filaments consist of protein subunits with three parts: the central rod domain, the N-terminal domain, and the C-terminal domain. The amino base of the N-terminal holds a positive electromotive potential in the highly charged E1 subdomain. The carboxyl acid of the C-terminal domain holds a negative potential in the highly charged E2 subdomain (shown in this illustration).
 The electrical properties and polarity of IF are consistent for all portions and all filaments. We will see later how MT filaments have different isozymes with diverse electrical properties that may be able to influence electrical flow in neurons. The electromotive potential in IF can serve as a bonding device for concatenating IF portions into a whole filament. It is unknown whether or not charged IF strands can perform any functions relevant to cognition.
The electrical properties and polarity of IF are consistent for all portions and all filaments. We will see later how MT filaments have different isozymes with diverse electrical properties that may be able to influence electrical flow in neurons. The electromotive potential in IF can serve as a bonding device for concatenating IF portions into a whole filament. It is unknown whether or not charged IF strands can perform any functions relevant to cognition.
 IF Arrangement
IF Arrangement
The rod domain consists of alpha-helical coiled coils of proteins. The subdomain organization for the central section of neurofilaments is not fully defined, although the termini of the end domains are better understood. The arrangement of oligomers in IF chains is also uncertain.
Neurofilaments consist of coiled coils: two IF chains combine to form a molecule; four make a 4-chain complex (also known as a protofilament); and eight 4-chain complexes make a segment of filament called a protofibril, which measures about 50 nm long and 4.5 nm across. The molecules are paired chains joined in parallel on the same axial register (a & b in the illustration).
The organization of the 4-chain complex of filaments is not yet established. If the end domains of the constituent coils are parallel (c), the 4-chain complex of filaments will be highly charged and polarized. If the end domains oppose (antiparallel – d below), the charges cancel each other and the filament will be nonpolar. A third possibility is that the chains form in staggered axial register so that end domains neither compound nor cancel each others’ charges (see below).

IF Termini
Determining the order of the termini may have important implications regarding possible information processing and storage capabilities. While all arrangements can transduce electrical potential, variations in the lattice organization of IF termini could serve variable cybernetic functions.
As Hameroff & Watt point out, charge and energy transfer, resonance, and long-range coherent interactions within proteins have already been described theoretically. This description bears a striking resemblance to parts of the hardware of a computer: “Oxidation/ reduction electron movement within proteins spatially fixed in an organelle or membrane has been compared to solid state electronics” [1982, p. 553]. Other suggested models include intraprotein electron dipole oscillations coupled to mechanical or conformational changes and cooperative processing with phase transitions, hysteresis and avalanche effects [ibid, p. 553].
It is reasonable to infer that IF are able to influence the electrical flow patterns within cells in ways analogous to the ways gates and transistors influence electrical flow patterns in integrated circuits. This concept will be revisited below.
 IF Protofibril
IF Protofibril

An IF protofibril actually consists of eight 4-chain complexes. This figure shows a possible arrangement for part of a single protofibril. If intermediate filaments possess both a structure similar to that suggested here (see figure at right) and variations in the structure (such as variable polarity) in terminal domains producing variable transductance, a strong argument for their participation in cybernetic activities could be made. Yet, as pointed out earlier and shown by Steinert and Roop [1988], many unknown factors may influence the formation of theories and models. Thus, all hypotheses about IF participation in cognitive functions must be considered totally theoretical.
IF Complexity
The IF gene family is complex. Because of their generally tissue-specific expression and the diversity of the properties of their end domains, Steinert & Roop say it “seems probable that IF are involved in specialized functions related to the differentiation state of the cell [1988, p. 619]. These specialized functions may include “mechanical coordination of the cytoskeleton of the cell or tissue, information transport, and signal transduction” [ibid].
Considering the complexity of IF – their polarity, their structure constituting an ability to participate in transduction of electromotive potential at different rates according to their terminal domain organization, and their great numbers in cell processes – they are potential candidates for an information storage or processing device in neurons.
IF may participate directly in information processing or they may interact with other parts of neurons to assist in learning or information acquisition. For example, we will soon be discussing possible cybernetic capabilities of MT. IF could have complex interactions with MT that differentiate the states of their components, thus contributing to learning.
IF help give and maintain neurons’ shapes, an aspect critical to their ability to form links with each other. This may be their most important cybernetic role. All cells have cytoskeletal filaments, but their shapes are often less critical to their roles in the body. Evidence supports both theories.
IF Properties
These four properties of intermediate filaments make them logical candidates for cybernetic components:
1) longevity–they are some of the more permanent neuron components. Longevity is important because humans retain knowledge throughout their entire lifespan.
2) electrical properties–they are able to transduce electrical potential. Electrical properties must support the flow of E/I (action potential).
3) location–they extend between neural points of reception and transmission. Location of cybernetic components must be in electrical flow pathways.
4) structure–they are filamentous and possibly possess multistable components. The structure must support the functions.
With the exception of longevity, all the characteristics favoring IF apply equally to microtubules. MT are slightly larger and slightly less stable than IF. Still, experts have suggested the possibility that MT perform cybernetic roles in neurons. The next post will address some of the possibilities for MT information-processing roles.
| Click below to look in each Understanding Context section |
|---|
| Intro | Context | 1 | Brains | 2 | Neurons | 3 | Neural Networks |
| 4 | Perception and Cognition | 5 | Fuzzy Logic | 6 | Language and Dialog | 7 | Cybernetic Models |
| 8 | Apps and Processes | 9 | The End of Code | Glossary | Bibliography |
SHARE THIS ARTICLE



