



14 Jan Visual Input Processing
 The visual cortex in the human brain is arguably the pattern after which most artificial neural networks were modeled: the flow of signals is directional through layers 1, 2 then 3; and large numbers of the cells are touched by the flow of action potentials through the system. The variations in the cells, however, contrasts with the artificial neural networks that all have identical nodes. In Section 2 of Understanding Context I have shown that all neurons are not identical, and may handle signals very differently.
The visual cortex in the human brain is arguably the pattern after which most artificial neural networks were modeled: the flow of signals is directional through layers 1, 2 then 3; and large numbers of the cells are touched by the flow of action potentials through the system. The variations in the cells, however, contrasts with the artificial neural networks that all have identical nodes. In Section 2 of Understanding Context I have shown that all neurons are not identical, and may handle signals very differently.
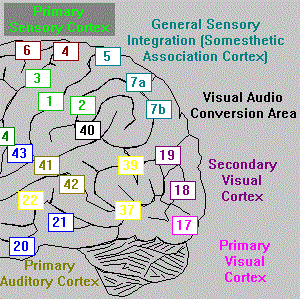
Signals coming in through the retina are received by a grid of cells that transmit the two dimensional patterns perceived to the visual cortex. They process successively through the visual layers I, II and III (Brodmann‘s areas 17, 18 and 19) then on to the conversion and integration areas. The conversion and integration areas are responsible for passing different types of visual patterns to different areas of the brain for further processing.
Simple Cells
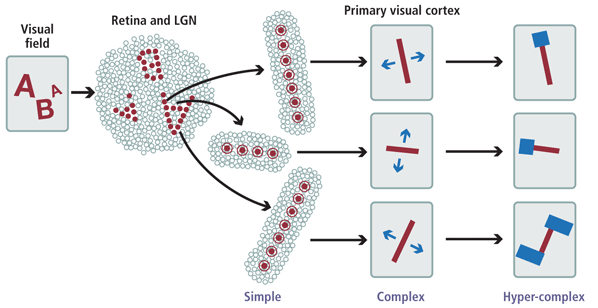
 Feature selective cells are usually described one of two ways: by classes or by where they reside on a scale of complexity. I will address the class divisions first. The features these types of cells detect are either line orientation or contrast, both of which provide parts of the visual input required to determine the shape of objects being viewed. Visual feature extractor cells in the striate cortex are categorized by their selectivity in response to stimuli from their associated receptive fields. A receptive field is the areas of the retina providing input for a cell.
Feature selective cells are usually described one of two ways: by classes or by where they reside on a scale of complexity. I will address the class divisions first. The features these types of cells detect are either line orientation or contrast, both of which provide parts of the visual input required to determine the shape of objects being viewed. Visual feature extractor cells in the striate cortex are categorized by their selectivity in response to stimuli from their associated receptive fields. A receptive field is the areas of the retina providing input for a cell.
The least feature selectivity is evidenced in simple cells. Their selectivity is most responsive to stationary input with spatially distinct “on” and “off” areas separated by parallel straight lines (Hubel & Wiesel, 1968, p. 217). Simple cells respond only to lines in the receptive field with a specific range of orientation. A substantial proportion of simple cells also exhibit color sensitivity. Detects stationary features fully within the cell’s receptive field. Detects stationary and moving features fully within the cell’s receptive field. Detects stationary and moving features partially but not fully within the receptive field.
| Understanding Context Cross-Reference |
|---|
| Click on these Links to other posts and glossary/bibliography references |
|
|
|
| Prior Post | Next Post |
| Harmonic Convergence of Light | Timing is Everything: Asynchronicity of E/I |
| Definitions | References |
| brain action potentials | Hubel 1968 Kuffler 1984 |
| feature cells | Hubel 1988 Martin Paré |
| sensory transduction | Vision Systems De Valois 1982 |
Complex Cells Selectivity to features anywhere in the receptive field is characteristic of complex cells. This flexibility enables them to respond to motion that does not activate simple cells. Tests using images (bars, slits and edges) on simple cells evoke responses throughout the receptive field in complex cells rather than in only one area, as in simple cells. Motion selectivity usually favors one direction, though motion of the same object in other directions evokes minimal response. Complex cells are ubiquitous in the visual cortex (Hubel & Wiesel, 1968, p. 218-219).
Hypercomplex Cells
Hypercomplex cells are typified by selectivity to features with corners or ends in the receptive field. They do not respond to lines or edges that extend completely through the receptive field and terminate elsewhere. The combination of simple, complex, and hypercomplex cells provides the visual system with the range of capabilities it needs to interpret any input within the visible spectrum. Continuum of Complexity The three distinct categories of cells – simple, complex, and hypercomplex – reside on a continuum based on conceptual complexity. This is not a physiological continuum, but one based on the differing specialized roles each plays in perceptual processing. As De Valois, et al., so relevantly state, “It is unfortunate that physiological investigations so often categorize cells into two or more different response types using qualitative methodology without presenting any quantitative evidence that the distribution of cells was in fact dichotomous rather than just varying uniformily (sic.) along a continuum” (De Valois, et al 1982, p. 549).
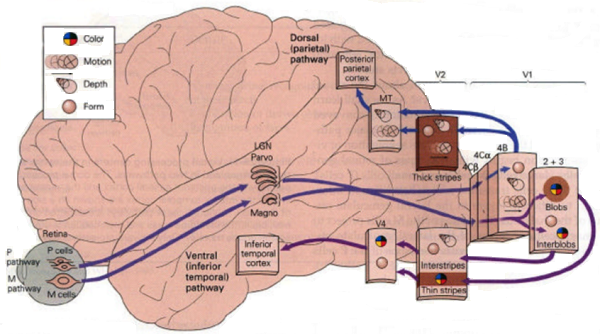
Image from Martin Paré at Queens University (brain.phgy.queensu.ca/pare/assets/Higher%20Processing%20handout.pdf)
Specialization of cells in the visual cortex to distinct and complementary functions is critical to the performance and quality of the visual input processing system in providing data for cognition. Now, let’s generalize the principle and suppose there are simple and complex cells performing various other cognitive functions in other parts of the brain, particularly areas involved in sensory perception. We have seen the structural differences between neurons such as purkinje cells and basket cells. Specialization seems to be the rule rather than the exception in almost all areas of the physiological constitution and functional organization of the nervous system. What can we infer from neural properties?
I’m planning a post to propose a set of parameters that may be inferred from what we know about neural properties. The most likely organization of data in the brain involves feature selectivity in perceptual processing centers and some binding process that enables association cells to link a set of simple features into a complex representation. I’m planning a post that extrapolates from these observations and shows what a maximum capacity neural storage model may be capable of holding.
| Click below to look in each Understanding Context section |
|---|
| Intro | Context | 1 | Brains | 2 | Neurons | 3 | Neural Networks |
| 4 | Perception and Cognition | 5 | Fuzzy Logic | 6 | Language and Dialog | 7 | Cybernetic Models |
| 8 | Apps and Processes | 9 | The End of Code | Glossary | Bibliography |
SHARE THIS ARTICLE



