



19 Jan Theories on Microtubules’ Function in Thought
Microtubule Functions
 Continuing to look inside neurons for the knowledge storage and processing mechanisms, let’s consider what Microtubules do. Microtubules serve many clearly identified functions. These functions include acting as structural members of the cytoskeleton, providing cellular orientation, and guiding membrane and cytoplasmic movements. In addition to these known functions, there is evidence that suggests neuronal MT may be associated with cell division and differentiation. They may also be associated with conscious perception, behavior, intelligence, and other cognitive processes [Hameroff & Watt, 1982].
Continuing to look inside neurons for the knowledge storage and processing mechanisms, let’s consider what Microtubules do. Microtubules serve many clearly identified functions. These functions include acting as structural members of the cytoskeleton, providing cellular orientation, and guiding membrane and cytoplasmic movements. In addition to these known functions, there is evidence that suggests neuronal MT may be associated with cell division and differentiation. They may also be associated with conscious perception, behavior, intelligence, and other cognitive processes [Hameroff & Watt, 1982].
For cybernetics-related functions, MT structure and MT organization or locations within neurons may play critical roles. Because of the structurally mediated motile capabilities of MT, it may be said to some extent that MT location is a result of MT structure (that is, MT are located throughout the cell body, axons, and dendrites because they have structural mechanisms that get them there).
| Understanding Context Cross-Reference |
|---|
| Click on these Links to other posts and glossary/bibliography references |
|
|
|
| Prior Post | Next Post |
| Microtubules (MT) | Gating in Human Reasoning |
| Definitions | References |
| Microtubules synaptic vesicle | Hameroff & Watt 1982 |
| cytoskeleton neurotransmitter | Linck 1989 |
| cybernetics synapse | Nadel 1989 |
The following functional matrix shows some of the known functions of microtubules.
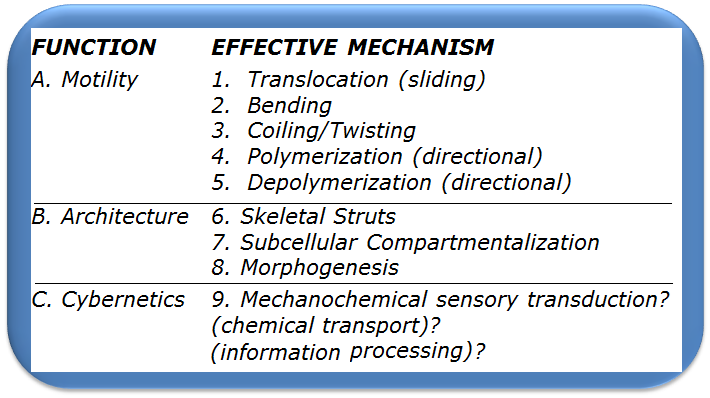
“?” signifies a theory with a shortage of scientific evidence. MT Functions [from Linck, 1989, p.14]
 Motility
Motility
Microtubules function includes motility: the function most frequently associated with MT in flagella and cilia. The organization of MT in flagella, such as those found in spermatozoa, is very regular. They slide and bend, causing the flagellum to wiggle. This wiggling provides locomotion. The primary motility agent is dynein. Flagellar dynein and IC, a dynein MAP found in neural MT, actively transport microtubules with a sliding motion.
In neurons, dynein-mediated sliding enables MT to act as rails along the length of axons from synapses to the soma, providing retrograde transport of subcellular structures, such as spent synaptic vesicles. Kinesin provides the motility for transportation from the soma of fresh vesicles and other materials. Another neural function of the motility of MT is in the neurite growth process. As a neurite grows toward the target point at which a new synapse will be formed, MTs provide the structural support, pushing and guiding the neurite through the maze. The actions of dynein and kinesin are coordinated by some “distributor” mechanism that is not fully understood.
 While MT provide flagella the ability to wiggle, their motility is also used in neurons:
While MT provide flagella the ability to wiggle, their motility is also used in neurons:
- to transport synaptic vesicles from the soma to the synapses and back; and
- to propel growing neurites.
Coiling, Twisting and Sliding
Coiling and twisting are documented MT activities, but they are not as well understood as dynein-mediated sliding and bending. Because of the mechanical complexity of coiling and twisting, these MT activities must be associated with conformational changes in the tubulin proteins of the MT cylinder. It is thought that these active changes either regulate interaction with force-generating enzymes or cause localized elasticity which could convert sliding to bending [Linck, 1989, p. 14].
While dynein is the agent in sliding, the agent in twisting may be actively changing molecular structures in the surface lattice of MT. This possibility highlights the complexity of these numerous subcellular structures and suggests possible roles MT might play in the growth and linking of neuron processes.
 The illustrations show neurite growth. The blue MT in the green neurite fiber represents many coordinated MT acting together to navigate through the crowded real estate of the brain to reach the target nerve fiber. This same capability is used for transporting bodies (organelles) within neurons. A synaptic vesicle with a neurotransmitter chemical (the yellow ball) must be transported from the soma where it is recharged to the synapse where it will be spent. MT perform this function by twisting and bending. MAPs help in this process.
The illustrations show neurite growth. The blue MT in the green neurite fiber represents many coordinated MT acting together to navigate through the crowded real estate of the brain to reach the target nerve fiber. This same capability is used for transporting bodies (organelles) within neurons. A synaptic vesicle with a neurotransmitter chemical (the yellow ball) must be transported from the soma where it is recharged to the synapse where it will be spent. MT perform this function by twisting and bending. MAPs help in this process.
Cytoskeleton Architecture
As mentioned in the posts on intermediate filaments (IF) and the post on cytoskeletal components, the structure of nerve processes is maintained by filaments. Actin microfilaments are by far the most numerous, while MT are the fewest. Both types play integral roles in providing for the morphological integrity of the cell. Microtubules’ role in mitosis or cell division may also be related to this architectural function.
MT form the centrioles that polarize in cells during mitosis. The spindles that form during mitosis and radiate from the centrioles are also MT. These play a role in determining which chromosomes go to which of the daughter cells. In embryonic development, the cell-division process and determination of chromosome inheritance makes differentiation possible. Thus, the overall architecture of the human body and the specific architecture of individual components relies, at least to some extent, on the MT’s structure-forming and preserving functions.
The architecture or structure of neurons has an impact on all the other areas pointed out in the main page of this section. Any attempt to divorce the structure of the neuron from its functionality would fail to capture the complexity and beauty of these incredible cells. The roles of MT in mitosis strengthen the contention that they may also be information bearers.
Microtubules in Cybernetics
Some researchers suggest that sensory transduction is accomplished by two principal mechanisms:
- Ciliary microtubules that generate and directly transmit chemical-electrical signals along their length; and
- MT that act indirectly by mechanically deforming membranes, thus inducing ion flow through channels [Linck, 1989, p. 15].
There is experimental evidence that the second mechanism, caused by bending of the tubule, enables MT to transmit stimuli to epithelial nerve cell junctions. It is possible that nerve membranes transduce the majority of the electrical potential involved in cognitive processing, but the likelihood of filaments acting as wires is equally plausible. Recent breakthroughs in understanding the in-vivo morphology of nerves (see “Neuron Shape”) seem to favor components in the cytoplasm as opposed to the long-held notion of membrane transduction as primary paths for currents.
Resistors, capacitors, transistors and other components affect the flow of electrical potential in man-made electrical circuits. Biological electrical circuits, particularly in the brain, may have similar capabilities. If they do, the possibility of single cells representing or remembering complex information is very real.
MT Lattice States
 The second principal mechanism of sensory transduction suggested by Linck [1989] is that MT act indirectly by mechanically deforming membranes, thus inducing ion flow through channels. These processes might explain apparent information functions within MT, including “cooperative resonance among spatially arrayed proteins, and propagated conformational changes along MT protofilaments” [Hameroff & Watt, 1982, p. 549]. Thus, the specific structure of the MT lattice and its component states may impact the level of transduction along MT strands in nerve processes. This could have the effect of selectively reducing the level of potential from multiple inputs prior to their reaching the axon and output terminals.
The second principal mechanism of sensory transduction suggested by Linck [1989] is that MT act indirectly by mechanically deforming membranes, thus inducing ion flow through channels. These processes might explain apparent information functions within MT, including “cooperative resonance among spatially arrayed proteins, and propagated conformational changes along MT protofilaments” [Hameroff & Watt, 1982, p. 549]. Thus, the specific structure of the MT lattice and its component states may impact the level of transduction along MT strands in nerve processes. This could have the effect of selectively reducing the level of potential from multiple inputs prior to their reaching the axon and output terminals.
Hameroff & Watt amplify what might be happening:
Propagated changes along the helical row axes as well as longitudinal protofilaments would imply that polymer subunits within cylindrical grid-like MT structure, connecting proteins and intracellular trabecular networks could provide programmable switching matrices for information transfer via boolean logic. Calcium dependent conformation states coupled to charge or energy quanta could be a medium of information transfer…with programming by genetic or environmental effects” [ibid, p. 550].
The idea of a microtubule or even a neuron being equivalent to a computer RAM chip in some functional way–such as the ability to store 1s and 0s to represent information–may seem a little far-fetched. Perhaps it is. Still, the analogy should not be discounted. We have already seen mechanisms that could act as gates in ways similar to the individual transistors in a RAM chip.
MT Isozymes
 With as many as 17 different isoforms or isozymes, the potential for MT to act as components of random access memory (RAM) or read-only memory (ROM) is structurally and functionally possible. The conceivable ability of MT to alter the electric potential flow within the cytoplasm is what makes this possible. In reality, though, the role of MT is probably limited to less complex perceptual data, such as image or audio features. The illustration on this page shows how a single MT in a dendrite could serve in several different roles. Multiple MTs could provide multiple selectivity to each dendrite and axon in a single neuron.
With as many as 17 different isoforms or isozymes, the potential for MT to act as components of random access memory (RAM) or read-only memory (ROM) is structurally and functionally possible. The conceivable ability of MT to alter the electric potential flow within the cytoplasm is what makes this possible. In reality, though, the role of MT is probably limited to less complex perceptual data, such as image or audio features. The illustration on this page shows how a single MT in a dendrite could serve in several different roles. Multiple MTs could provide multiple selectivity to each dendrite and axon in a single neuron.
In order to understand the mechanism of information processing suggested by this theory, we must examine the mechanisms and modes of information processing between neurons. In the next section (Volume 3), connections between neurons will be described. We will also examine processes that mediate the spreading of excitation and inhibition, collectively called action potential, to discover what processes could accommodate a model of explicit information storage in neurons.
MT and Sensory Input
In addition to structural properties, there is also functional evidence that MTs play an important role in information-processing, particularly in sensory transduction. Certain mutations have been found in which organisms have developed one extra protofilament in some neural microtubules. This was discovered in nematodes; the result was that chemoreceptor-pattern processing was disabled. This association between MT and sensory-input pattern processing provides a clue to information-processing possibilities in neurons. Although the evidence is remote and too scant to confirm the theory, there are grounds for speculation on how MT might perform cybernetic functions.
Individually, MT have a structure that is complex enough to consider them as information-processing mechanisms. In addition, bridges between MT add to the complexity – as well as the information capacity and processing potential. Add to these relationships the presence of other cytoskeletal members (IF and microfilaments), and the possibility for complex information storage mechanisms within each neuron becomes quite plausible.
 Mutations and pathologies often give good clues to the functions of some parts of the brain and its components. Just as aphasia (receptive and expressive) tell us about the functions of Broca’s and Wernicke’s areas, researchers are studying nematodes with anomalous MT may give us evidence that will help us better understand MT. Hopefully, there won’t be too many things we have to break in order to find out what MT are designed to do!
Mutations and pathologies often give good clues to the functions of some parts of the brain and its components. Just as aphasia (receptive and expressive) tell us about the functions of Broca’s and Wernicke’s areas, researchers are studying nematodes with anomalous MT may give us evidence that will help us better understand MT. Hopefully, there won’t be too many things we have to break in order to find out what MT are designed to do!
Summary
Most of the discussion in this section of the blog has focused on the potential of neurons’ bone structure (cytoskeletal members) to participate in cognitive activities. A deep examination of other components of cells may turn up similar structural and/or functional properties that suggest participation in perceptual, memory or thought processes. So much is still unknown! The best we can do is zero in on areas that appear likely to be involved in the electrochemical processes of action potential flow to try to discover what in heaven’s name they are doing.
In the blog posts on “Networking”, we will look at action potential flow. We will describe some of the critical processes associated with determining the intensity of excitation and inhibition, the duration of salient electrical activity, and the frequency of recurring action potential spikes. We began at the macroscopic level of the brain in Section 1 Brains), then moved down to the level of the neuron in Section 2 (Neurons). Section 3 (Neural Networks) deals mostly with the synapse: that tiny gap over which the sparks of thought ignite the flame of human reason.
| Click below to look in each Understanding Context section |
|---|
SHARE THIS ARTICLE



